
Getting Closer to Real Patient Biology
Advanced preclinical platforms that mimic native tissue complexity, especially those that inherently include epithelial, stromal, and immune compartments are reshaping how biopharma evaluates new therapies. These models are more than just tools for testing—they provide insight into disease mechanisms, stratify patient responses, and reduce the risks of clinical failure.
Four Levers That Matter in Preclinical Strategy
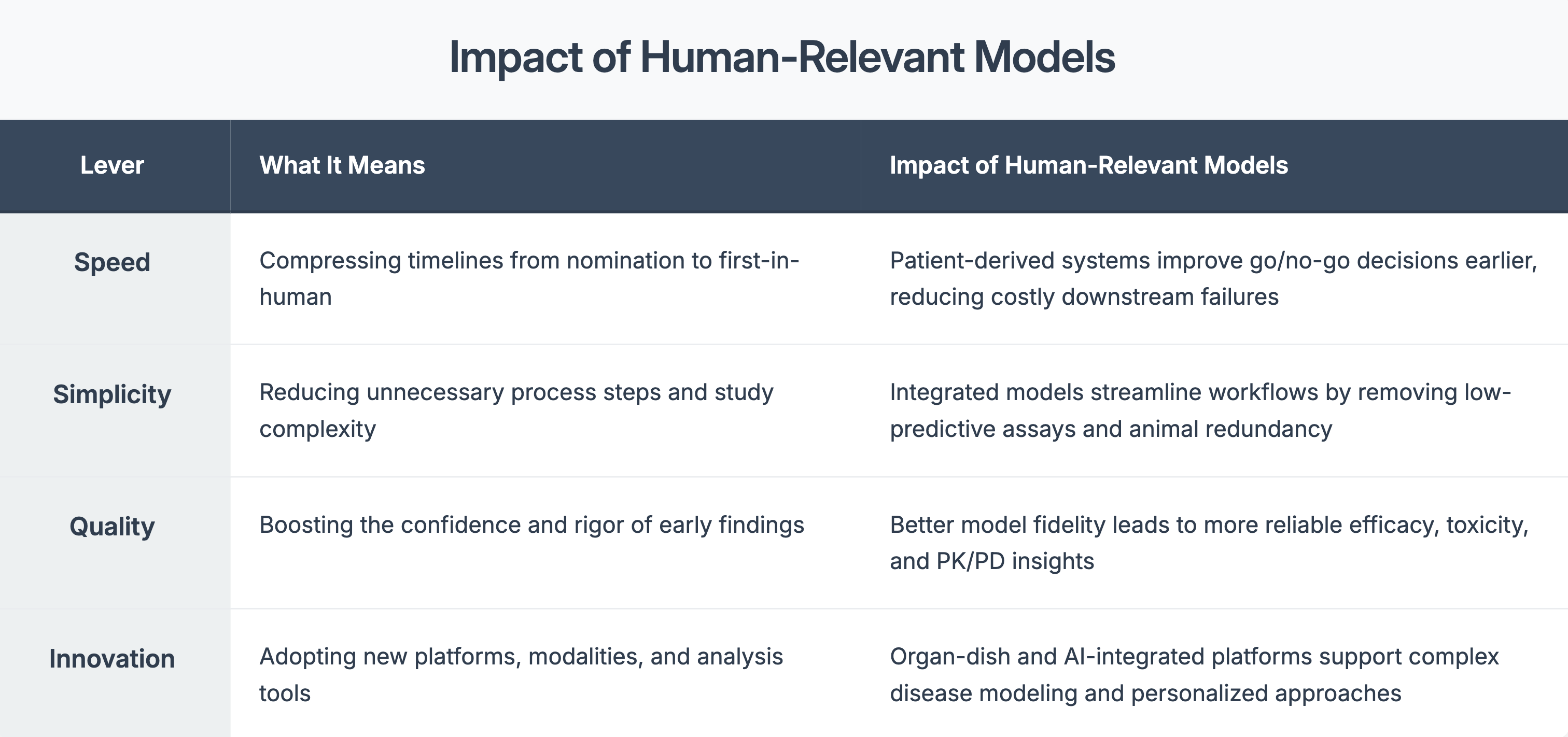
When used early, they can:
• Flag toxicity profile before trials begin
• Reveal differences in drug response across patient subtypes
• Guide dose optimization with more confidence
- Support regulatory filings with more compelling data
References
- 1. Agrawal G, Bader F, Günthner J, Wurzer S. Fast to first-in-human: Getting new medicines to patients more quickly. McKinsey & Company; 2023. 2. Harvard Wyss Institute. Beyond animals: Revolutionizing drug discovery with human-relevant models. 2024. 3. Charles River. Leveraging advanced technology to expedite oncology R&D. 2024. 4. Regulatory Scoping Review. Clinical relevance in preclinical model selection. PharmTech & PubMed; 2024. 5. Frontiers in Toxicology. AI-powered digital toxicology platforms. 2024. 6. Wikipedia. 3D Cell Culture & Patient-Derived Xenografts. Accessed 2024.